キンボスポンディルス
| キンボスポンディルス | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||
| 地質時代 | ||||||||||||||||||||||||
| 前期三畳紀オレネキアン - 後期三畳紀アニシアン | ||||||||||||||||||||||||
| 分類 | ||||||||||||||||||||||||
| ||||||||||||||||||||||||
| 学名 | ||||||||||||||||||||||||
| Cymbospondylus Leidy, 1868 | ||||||||||||||||||||||||
| シノニム | ||||||||||||||||||||||||
| 種 | ||||||||||||||||||||||||
キンボスポンディルス(学名:Cymbospondylus)は、三畳紀に生息した魚竜目に属する大型海棲爬虫類の属。体サイズは種によるが、全長10メートルに達する種も報告されている[1]。1868年に記載されたアメリカ合衆国ネバダ州産の部分的な化石が最初に発見され、その後1900年代初頭に全身骨格が発見された[1]。長い尾を左右に振って遊泳し、海棲動物を捕食していたとされる[1]。
研究史
発見と同定
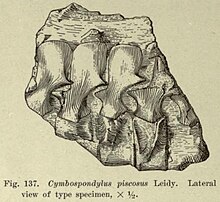
1868年、アメリカ合衆国の古生物学者ジョゼフ・ライディは中部三畳系から産出した魚竜の2新属を記載した。記載に際して基づいた標本は同国ネバダ州の複数の産地から回収された椎骨の化石であり、いずれも地質学者ジョサイア・ホイットニー(英語版)を介して提供されたものであった。2属のうち1つがキンボスポンディルスであり、ライディは本属に2種を分類した。第一の種はC. piscosusで、ライディは様々な山脈で発見された脊椎に基づいて命名した[2]。C. piscosusのホロタイプ標本はネバダ州オースティン(英語版)北西部にてNew Pass Rangeで発見された不完全な5個の胴椎を含むブロックである[3][4][5][6][注釈 1]。ライディは他の2個の標本を本種に分類しており、1個はHumboldt Rangeのスターキャニオンで発見された一連の8個の尾椎で、もう1個はToiyabe Rangeで発見された同じく尾椎と推測される1個の椎骨である。第二の種C. petrinusはHumboldt Rangeで発見された5個の大型の胴椎に基づいて命名された[2]。属名Cymbospondylusは古代ギリシア語で「カップ」を意味するκύμβη (kymbē)と「椎骨」を意味するσπόνδυλος (spondylos)に由来し、「カップ状の椎骨」という意味になる。これは骨格のうち椎骨の明らかな形状を反映したものである[7]。1868年の論文ではタイプ種が指定されていなかったため、1902年にジョン・キャンベル・メリアム(英語版)がC. piscosusをタイプ種に指定した[3]。本種はライディによる本属の公式な記載において最初に命名されたものである[2]。
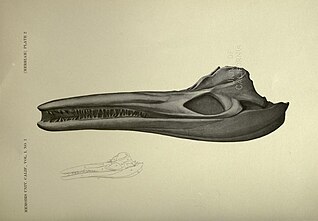

1901年から1907年の間に、カリフォルニア大学は三畳紀の魚竜を可能な限り多く回収してより詳細な調査をするため、合衆国内の様々な場所へ10回の発掘遠征を実施した。これらの発掘遠征はメリアムによって主導され、ほぼ全てがアニー・モンタギュー・アレキサンダー(英語版)による資金援助を受けた。この調査により最終的に50個を超える標本が回収され、それぞれが骨格のかなりの部分を保存していた[8]。これらの化石の中にはUCMP 9950に指定されたほぼ完全な骨格を含め非常に保存の良好なC. petrinusの化石が複数あり、その全てがライディから言及された原産地で発見された。研究で言及された他の魚竜と同様に、メリアムは新たに知られた化石に基づいてキンボスポンディルスをより詳細に記載した[9]。メリアムによるC. petrinusの解剖学的記載はいまだ有効とみなされており、キンボスポンディルスの後続研究における比較解剖学にも用いられ、本種に妥当性をもたらしている[5][10][6][11][12]。
C. petrinusと異なり、タイプ種C. piscosusの追加標本は発見されていない。メリアムの研究では、体サイズが小型であることとホロタイプ標本の椎骨に存在する規則的な窪んだ表面に基づいてC. piscosusを識別可能とされた[3][13]。しかしその後C. piscosusの有効性に疑問が投げかけられ、複数の研究者が当該化石の疑わしい性質に言及した[5]。McGowan and Motani (2003)はC. piscosusの椎骨が種の有効性を証明する標徴形質を示さないと指摘した。タイプ種が疑問名である場合属そのものも疑問名になるため、この事態は分類学的問題を生じることとなった。この問題を解消するため、マッゴーワンと藻谷はC. petrinusの最も完全な標本 UCMP 9950 をC. piscosusのネオタイプに指定できることを示唆した[14]。しかし、動物命名法国際審議会への正式な審判請求が行われていないため、提案されているネオタイプに対する学名C. petrinusは追って通知があるまで保持される[6]。Wolniewicz (2017)はメリアムにより記載された追加の化石をC. piscosusに分類し、C. petrinusとC. piscosusとをシノニムとした[15]。しかしこの提案は博士論文に留まっており、国際動物命名規約条8により公表された著作物に掲載されたものと見なされないため、正式に有効でない[16][17]。従って、C. piscosusは本属のタイプ種であるものの、本属の大半の記載には含まれていない[11][12]。
認められた種
1927年、スイスのサン・ジョルジョ山の産地にて、ベザーノ層で大型の魚竜の骨格が発見された。この化石はベルンハルト・ペイヤー(英語版)により1944年に言及された[18]。その20年後、Kuhn-Schnyder (1964)は当該標本の写真を公開し、当時北アメリカからしか化石が産出していなかったキンボスポンディルスと親和性を共有していることを示唆した[19]。件の標本はPIMUZ T 4351に指定された後、ポール・マーティン・サンダー(ドイツ語版)により1989年にC. buchseriとして記載・命名され、ヨーロッパにおけるキンボスポンディルスの存在が確認された。種小名buchseriは最初の主な専門的業績として1931年にホロタイプ標本のプレパレーションを行ったチューリッヒ大学古生物学博物館のメンバーであるFritz Buchserへの献名である[5]。

C. petrinusはネバダ州から化石が産出しているうちで有効な唯一の種であったが、21世紀初、Fröbisch et al. (2006)がネバダ州のAugusta Mountainsで発見された不完全な骨格標本FMNH PR 2251に基づいてC. nichollsiを記載した。この化石は当初C. petrinusの新標本の発見に期待されて発掘されたものであったが、解剖学的差異が多いことから別種として確立されることになった。本種の種小名は三畳紀の魚竜研究に多大な貢献を残した、三畳紀の海棲爬虫類の専門家であるアメリカ系カナダ人の古生物学者エリザベス・ニコルズ(英語版)への献名である[6][7]。この記載において、FröbischらはC. petrinusに分類されたほぼ完全な骨格UCMP 9913[注釈 2]が実際には形質の類似性からC. nichollsiに属する可能性があるとした。しかし、当該標本が科学文献であまり扱われていないため、Fröbischらは本標本がC. petrinusの種内変異を示すかどうかが判断できずより詳細な記載が必要であると判断した[6]。キンボスポンディルスの後続研究においてUCMP 9913はFröbischらの示唆した可能性と異なりC. petrinusに分類されているが[21]、顕著な差異も同時に言及されている[11]。Wolniewicz (2017)はC. nichollsiを解剖学的に再記載しこれをC. piscosusのジュニアシノニムとしたが[22]、この見解は共有されておらずその後の後続研究で本種は有効な種として扱われている[11][12][21]。
2011年には新しいキンボスポンディルスの化石がAugusta Mountainsで発見され、3年後の2014年に発掘された。この部分的な骨格は2013年に同時代の大型魚竜タラットアルコンの記載で言及され[23]、2018年の組織学的研究に際しても言及され分析対象となった[24]。本標本 LACM DI 158109 はKlein et al. (2020)でC. duelferiのホロタイプとして指定された。本種の種小名は中生代の海棲爬虫類の調査に貢献した化石プレパレーターのOlaf Dülferへの献名である[11][7]。

1998年にサンダーはAugusta Mountainsでキンボスポンディルスの別の標本を発見し、2014年から2015年にかけて仲間と発掘を行った[21]。プレパレーションの後、本標本LACM DI 157871は大型の完全な頭骨・複数の頸椎・右上腕骨と断片的な肩帯からなることが判明した。C. duelferiの同定から1年後、本標本はSander et al. (2021)によりキンボスポンディルス属のさらなる新種として同定された[12][21]。本種C. youngorumの種小名は発掘プロジェクトを資金面で支えた[25]Tom Young と Bonda Youngへの献名である[12]。
本属に属する可能性のある種
より部分的なキンボスポンディルスの標本はヨーロッパの様々な地層で他にも多数発見されているが、具体的な分類は確定しておらず、科学文献ではキンボスポンディルス属未定種(Cymbospondylus sp.)として扱われている。これらの標本のうち3個はアイダホ州(アメリカ合衆国)、2個はスヴァールバル諸島(ノルウェー)から産出しており、共に下部三畳系のオレネキアン階にあたる。このため、これらの化石はキンボスポンディルス属のうち最古の標本群となっている[26][27][28]。
キンボスポンディルス属に潜在的に含まれる可能性のある標本を以下に列挙する。
- イタリアのMonte Seceda(イタリア語版)から産出した魚竜の前半身を保存した部分的骨格についてKuhn-Schnyder (1980)が記載した。記載された標本は当初シャスタサウルス属未定種(Shastasaurus sp.)に分類されていたが[29]、サンダーが1989年にC. buchseriを記載した際に本標本もキンボスポンディルス属に再分類された[5]。この分類が現代も認められている[30]。
- ノルウェーのスヴァールバル諸島に位置するスピッツベルゲン島の別々の産地から産出した2個の標本をSander (1992)が記載した。論文中で最初に記載された標本PIMUZ A/III 496は1961年にBotneheia(英語版)で発見された標本であり、より保存が良好で、肋骨と関節した17個の椎骨からなる。2番目の標本PIMUZ A/III 554 および 555は2個の単離した椎骨からなり、Wichebukta(英語版)という湾で発見されたものである[31]。
- アメリカ合衆国アイダホ州のThaynes FormationでH・グレゴリー・マクドナルドが1985年に発見した多数のキンボスポンディルスの化石について、Massare and Callaway (1994)が言及した[26]。
- Rieppel and Vecchia (2001)はイタリアのフォルニ・ディ・ソットで発見された三畳紀の海棲爬虫類を一覧化しており、その中には間違いなくキンボスポンディルスとされる2個の化石も含まれている。これらのコレクションのうち1個目の化石は1個の椎骨・1本の神経棘・3個の肋骨断片であり、2個目の化石は2個の単離した椎骨からなる[32]。
- イタリアのピアッツァ・ブレンバーナで発見された多少部分的なキンボスポンディルスの4個の椎骨について、Balini and Renesto (2012)はMCSNB 11689 A, B, C, および Dの標本番号の下で言及した[30]。
- 2013年と2018年には、スヴァールバル諸島のVikinghøgda層Vendomdalen部層で産出した多数の椎骨が属レベルで同定された[33][27][28]。



かつて分類された種
キンボスポンディルス属には有効かつ固有派生形質を持つ種が多数分類されている。しかし幾つかの種はその分類史を通し、別属に再分類されるか、既存の種のシノニムとされるか、あるいは疑問名とされてきた[6]。Leidy (1968)はキンボスポンディルスの記載の際、Humboldt Rangeのスターキャニオンで発見された尾椎の断片に基づいてChonespondylus grandisを命名した[2]。本種の学名のうち属名は「漏斗」を意味する古代ギリシア語のχοάνη (khoánē, "funnel") と「椎骨」を意味するσπόνδυλος (spondylos)に由来し、種小名はラテン語で「大型な」「幅広な」を意味するgrandisに由来する[7]。Merriam (1902)はライディの発見を一覧化したが、ここでChonespondylusをキンボスポンディルス属から区別する十分な形質が存在しないとし、前者を後者のシノニムとしてC. (?) grandisとして扱った[34]。その後、C. petrinusの非常に完全な新しい化石の発見に伴い、Merriam (1908)はC. (?) grandisをC. petrinusのシノニムとした[35][6] 。
Hulke (1873)はイクチオサウルス属の種I. polarisを記載し、ノルウェーのスピッツベルゲン島に位置するイスフィヨルデン(英語版)で発見された2組の肋骨断片と椎骨とに基づいて命名した[36]。1902年にロシアの古生物学者Nikolajewitch Yakowlewは本種をシャスタサウルス属に再分類し、単離した1個の椎骨を本分類群のものに分類した[37]。Merriam (1908)は本種をキンボスポンディルス属に戻して学名をC. (?) polarisとしたが、既知の化石が乏しいために真に分類を決定することができないとした[38]。Wiman (1910)が本種をP. polarisとしてペソサウルス(英語版)属に再分類して以降[39]、本種はペソサウルス属の種として扱われている。本分類群は20世紀末に出版された研究によれば疑問名とされることもあるが、McGowan and Motani (2003)によれば未確定種(英語版)であり、本種に分類される多数の化石が再び有効になる可能性のある調査中の分類群とされている[40]。
Merrian (1908)はC. petrinusと同一産地から産出した標本に基づき、本属に2新種を設立した。1種目は後肢を構成する化石から命名されたC. nevadanusであり、Merrian (1908)は複数の骨のプロポーションが異なることや本標本の骨の方が大型であることから、本標本をC. petrinusと区別した[41]。しかし、C. nevadanusの化石は種の有効性を支持するには標徴形質が不十分であり、McGowan and Motani (2003)により未確定種(英語版)と判断された[42][6]。Merrian (1908)により設立された2種目はC. natansで、単離した上腕骨・尺骨・橈骨・中手骨および一連の尾椎に基づいて命名された。Merrian (1908)はこれらの骨がミクソサウルス属との類似性を持つことを指摘し[43]、後にMerriam (1911)でM (?) natansに改名することになった[44]。20世紀の大部分においてM (?) natansはミクソサウルス属の有効な種と見なされていたが、Nicholls et al. (1999)によりM. nordenskioeldiiのシノニムとされた[45]。M. nordenskioeldii自体は2005年以降疑問名と考えられているが[46]、関係する化石は依然としてミクソサウルス科(英語版)に分類されており、もはやキンボスポンディルス属には分類されない[6]。
Huene (1916)はドイツの魚竜をレビューし、バーデン=ヴュルテンベルク州のラウフェンブルグ(英語版)のMuschelkalkで発見された化石に基づき、キンボスポンディルス属に2種を設立した。1種目のC. germanicusは椎骨を伴った1本の肋骨や基後頭骨(英語版)に基づいて命名された[47]。直後に本種の有効性はBroili (1916)により疑問視され、独立した種として判断できるだけの特徴が存在しないと指摘された[48][6]。加えてこの化石は有効な種として判断されるには保存が劣悪であり[5]、これらを踏まえて疑問名とされた[49][50]。

Li and You (2002)は中華人民共和国貴州省のXiaowa層から発見された1個の完全な頭骨に基づいて新種C. asiaticusを命名した[51]。2006年に出版されたC. nichollsiの正式な記載では、著者らは本種をキンボスポンディルス属に分類することに疑問を呈した。彼らはC. nichollsiが当時知られていたキンボスポンディルス属の種であるC. petrinusとC. buchseriおよびC. nichollsiとの顕著な共通点を示さないことを指摘し、実際にはGuizhouichthyosaurus tangaeのシノニムであることを示唆した[6]。Guizhouichthyosaurusが同一の層から発見された後、Fröbischらによるシノニム化の提案はShang and Li (2009)により受容されたが、Guizhouichthyosaurusがシャスタサウルス属に類似することから本種はShang and Li (2009)によりS. tangaeへ再分類されることとなった[52]。この分類変更はその後も争点となり、Maisch (2010)が暫定的にGuizhouichthyosaurusを独立属に分類した後[53]、Sander et al. (2011)もGuizhouichthyosaurusを独立属とし[54]、Shang and Liがシャスタサウルスとのシノニムとした[55]。しかし、公表された数多の系統解析と形態解析では、Guizhouichthyosaurusはシャスタサウルスと異なる属に位置付けられている[56][57][58][59]。
特徴

他の魚竜と同様に、キンボスポンディルスは長く薄い吻部・大型の眼窩・下側を椎骨に支持された尾鰭を持つ。魚竜は表面的にイルカに類似しており、後肢よりも長い前肢の鰭を持つが、パルヴィペルヴィア類を除く古いグループでは背鰭が存在しないかあるいは比較的発達していない[60]。大半の三畳紀の魚竜と同様に、キンボスポンディルスの解剖学的構造は細長く、長く伸びた胴部とあまり目立たない長い尾を有する[61][62][30][63]。
体サイズ

キンボスポンディルスの体サイズと体重は種間で大きく異なる[6][11][28]。C. petrinusの体サイズ推定は完全な骨格標本UCMP 9950が発見されたことなどによって1908年以降比較的変化している[64]。メリアムはUCMP 9947とUCMP 9950に基づいてC. petrinusの全長を9.1メートルを超過するものと推定した[65][5][6]。Klein et al. (2020)はC. petrinusの推定値をより大きくし、全長を9.3メートル、頭骨長を1.16メートルとした[11]。Sander et al. (2021)はC. petrinusの全長を12.56メートル、体重を5.7トンと推定したが、頭骨長の推定値については先行研究と同様であった[21]。この推定値の起源は、UCMP 9947とUCMP 9950に基づいてC. petrinusのサイズについてさらなる改訂を行い、2つを組み合わせて合計で約10メートルの値が得られることによるとSander et al. (2024)で説明されている。標本UCMP 9947は後側の尾椎が複数失われており、そのため全長が増加して11メートルから12メートルを超過しても不合理ではない[28]。
C. youngorumのホロタイプ標本は頭骨長が2メートル近くに達し、上腕骨が魚竜の記録の中で2番目に大きいものになっている。最大の全長推定値は17.65メートルに達し、体重推定値は44.7トンに達する[注釈 3]。この推定によりC. youngourumはイクチオティタン(英語版)に次ぐ最大級の魚竜の一つになっただけでなく、中期三畳紀における最大級の動物の一つとなった[12][28][66]。
C. nichollsiのホロタイプ標本が骨格の大部分を保存しているものの、頭骨が半分しか保存されていないため、本種の体サイズ推定は複雑である。しかし、C. buchseriやC. petrinusとの比較に基づき、頭骨長が97センチメートル、全長が7.6メートル[6][11][28]、体重が3トンと推定されている[21]。C. buchseriのホロタイプの推定値は僅かにこれよりも小さく、全長約5 - 5.5メートル、頭骨長68センチメートルと推定されているが、体重の推定値は公表されていない[5][59][28]。C. duelferiは本属において既知の範囲内で最小の種であり、頭骨長が65 - 70センチメートルであり、正式に記載された研究で全長推定値が4.3メートルである[11]。Sander et al. (2021)による彩度の推定では、全長約5メートル、体重520キログラムの推定値が得られた[21][28]。
種が特定されていない個体についても体サイズの推定値が与えられている。C. youngorumに適用されたものと同様の推定方法により、Vikinghøgda層から産出した標本IGPB R660は全長7.5 - 9.5メートルと推定されており、前期三畳紀で既知の範囲内における最大級の魚竜の一つとなった[28]。
頭部

他の魚竜と同様に、キンボスポンディルスの吻部は長く伸びた円錐形であり、前上顎骨と鼻骨が最大の骨となっている[65][6][11][21]。鼻骨は遥かに後側へ伸びており[67][5][11]、前頭骨と共に側頭窓の前縁へ達する[6][11]。眼窩は楕円形から卵形であり、頭骨の大きさに比較してプロポーション的に小型である[65][5][6][11]。上側頭窓の形状はいくつかの種間で異なっており、C. nichollsiとC. duelferiで楕円形、C. petrinusで三角形をなす[11]。強膜輪を構成する強膜小骨の数も種間で異なり、C. duelferiで12個[11]、C. buchseriで13個[5]、C. nichollsiで14個[6]、C. petrinusで14個から18個である[68]。C. youngorumにも強膜輪が存在するが、強膜小骨の個数については言及が無い[21]。矢状稜(英語版)はC. duelferiとC. nichollsiにおいて低く、C. petrinusで明らかに高く[注釈 4][11]、C. youngorumで全く存在しない[21]。キンボスポンディルスの後頭顆(英語版)は窪んでいる[6][11]。他の魚竜と同様に、キンボスポンディルスの下顎は長く薄く伸びており[69]、頭骨の後方まで後側に伸びている[6][11]。下顎を構成する主要な骨である歯骨は眼窩の中央付近と同位置まで伸びる[69][6][5][11]。上角骨は薄くなりながら後関節突起まで伸びる。角骨は下顎の外側の腹側部を構成し、縫合線を介して上角骨と接する[6][11][21]。
キンボスポンディルスの歯列は槽生歯であり、歯根が顎骨に深く固定されている。しかし全ての種が同様に堅牢な構造を共通して持つわけでなく、C. petrinusの歯が歯槽の底部で癒合するように見える一方[70]、C. duelferiの歯は浅い歯槽に埋め込まれており[11]、C. youngorumの歯は他の魚竜に見られない特徴である[12]厚い骨の基部が歯に接する[21]。歯は同歯性であり、円錐形で稜を持ち突出した形態の歯が共通している。歯は歯冠の基部から頂点に伸びる長軸方向の溝を有する[11][21]。種ごとの歯の合計本数の差異は特定困難である。これは、特定の化石の保存状態が正式な評価を妨げているためであり、具体的にはC. buchseriで保存が悪く[5]、C. nichollsiで全く確認されていない[6]。歯の本数を多少なりとも明確に推定可能な種は3種のみである。C. duelferiは下顎の歯の本数が不明であるが、上顎の歯の本数が21本を上回る[21]。C. petrinusは上顎と下顎に30本から35本の歯を持ち[71]、C. youngorumは上顎に43本の歯と下顎に31本を上回る歯を持つ[21]。
分類
系統
魚竜類におけるキンボスポンディルスの正確な位置付けは不明であり、研究ごとに結果が異なる。場合によってミクソサウルス科よりも基盤的な位置に置かれることもあれば、逆に派生的な位置に配置される場合もある[72]。ただし、いずれにせよキンボスポンディルスが魚竜の分岐群において基盤的な位置にあることは合意されている。初期の系統解析においてキンボスポンディルスはシャスタサウルス科に配置された[73]。Bindellini et al. (2021)では魚竜の最も基盤的な位置に置かれ、ミクソサウルス科とシャスタサウルス科を含む分岐群であるHueneosauriaのより派生的なメンバーの外側に位置付けられた[59]。Klein et al. (2021)はC. duelferiの記載に際してネバダ州のFossil Hill部層から産出した全ての種が互いに分岐群を形成する樹形を再現した[11]。このネバダ州の分岐群についてはC. youngorumの記載によりさらに情報が加えられ、C. youngorumが最も派生的な種でヨーロッパのC. buchseriが最も基盤的な種に位置付けられた。Bindellini et al. (2021)における解析と同様に、シャスタサウルス科とミクソサウルス科はより派生的な魚として配置された[12]。
先駆けて行われた多くの解析と同様に、キンボスポンディルス属のタイプ種C. piscosusはKlein et al. (2021)の解析のデータセットに含まれなかった[12]。2020年の研究では、C. nichollsiの頭骨の形態がレビューされ、C. petrinusの形態と類似するものの側頭窓の形状から別種として区別できるため有効な種と判断された[11]。
以下のクラドグラムはSander et al. (2021)に基づき、魚竜類におけるキンボスポンディルスの位置づけを示す[12][21]。
| Ichthyosauromorpha |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
進化
魚竜は約2億4800万年前から約9000万年前にかけての前期三畳紀末から後期白亜紀初頭まで、中生代の大部分の時代において繁栄した海棲爬虫類の主要なグループの一つである[60]。キンボスポンディルスはこの魚竜の歴史において早期に出現しており、これまでに同定された魚竜のうち最古級の例の一つである。しかし初期の魚竜にも拘わらず、本属は進化を通して特定の魚竜が急速に大型化したことを示唆している。事実、カートリンカスのようなIchthyosauriformes(魚竜とその祖先や近縁系統を含むグループ)の最古の代表例が頭骨長5.5センチメートル程度である一方、既知の範囲内でキンボスポンディルス属の最大の種であるC. youngorumは頭骨長約2メートルに達しており、さらにこれら2分類群の生息年代は僅か250万年しか離れていない。類似する四肢動物である鯨類を見てみると、頭骨幅12.7センチメートルのパキケトゥスと頭骨幅60センチメートルのバシロサウルスとの間では約1400万年の開きがある。ハクジラ類では、頭蓋骨幅23.8センチメートルのシモケトゥス(英語版)と頭蓋骨幅約2メートルのリヴィアタンとの間で2500万年を超過する隔たりがある。魚竜に見られる急速な大型化は、ペルム紀末の大量絶滅後のコノドントとアンモナイトの急速な多様化により促進された可能性がある。なお魚竜はその後大型の目を進化の過程で獲得することになるが、こうした発達した目は食糧源をより精度良く特定可能になることから、魚竜の体サイズの顕著な現象に繋がったとされる[12]。
古生物学
Massare and Callaway (1990)はキンボスポンディルスを含む三畳紀の魚竜が待ち伏せ型捕食者であった可能性を提唱した。彼らの主張によれば、後世の魚竜の持つ三日月型の尾鰭を欠いた側方に平たい尾が波打つ遊泳様式に適しており、長い頸部と胴部が水中で抗力を生み出したと考えられる。彼らは、初期の魚竜に見られる長い体が柔軟な体波打つ遊泳様式を支持する構造を持ち、強力な尾が一気に速度を出していたとし、いずれも待ち伏せ型の捕食への適応の可能性があるとした。また彼らは、キンボスポンディルスをコンパクトでイルカ型の体を持ち追跡型捕食者に適した連続的遊泳を行うジュラ紀の魚竜と比較し、対照的とした[74]。
スイスから産出した中型のCymbospondylus buchseriは直接的な食餌の証拠が得られており、胃内容物が鞘形亜綱(英語版)の頭足類のものである腕の鉤のみで構成されている状態で発見されている。しかし、生前最後の食餌は典型的な食餌を正確に反映していない可能性があり、これはC. buchseriがより大型の獲物を捕食した可能性を排除するものではない。Bindellini et al. (2021)はC. buchseriがより緩慢な食餌サイクルやより高い咬合力を伴うより強力な採食戦略をとっていた可能性に触れ、その根拠として頑強な吻部を挙げた。ベザーノ層では、キンボスポンディルスはより華奢な頭骨を持つベサノサウルス(英語版)やミクソサウルス類といった他のより小型の魚竜と共存していたとされる。C. buchseriがより大型の獲物を捕食したかはともかく、3つの分類群は全て異なる狩猟戦略や獲物の嗜好に対する明確な適応を示している。ただし、その生態学的詳細は完全には明らかにされていない[59]。

鈍く円錐形の歯と長く伸びた吻部から、C. youngorumは頭足類や魚類を捕食するジェネラリストであったと推測される。しかし、C. buchseriと同様に、Sander et al. (2021)はC. youngorumが同地域の同属他種を含めて大型の脊椎動物をも捕食していた可能性を指摘した[12]。
Bindellini et al. (2021)は、キンボスポンディルスの絶滅がシャスタサウルス科の多様性に寄与した可能性を指摘した。中国のカーニアン階からは生態学的に異なる3種類のシャスタサウルス科が発見されているが、その一方でキンボスポンディルス科は既に絶滅しており化石記録が無い[59]。
古環境
北アメリカ

北アメリカから産出したキンボスポンディルスの種は、いずれもネバダ州に分布するスターピーク層群(英語版)の2つの地層から化石が産出している。C. petrinus、C. nichollsi、C. duelferi、C. youngorumはFavret層から産出しているが、最初の2種はPrida層で同定された本属の数少ない既知の例でもある[6][11][12]。これらの2つの地層はFossil Hill部層として知られる単一の部層で繋がっている。Prida層においてこの部層の露頭はHumboldt Rangeの西部に位置し、Favret層ではAugusta Mountainsに位置する[75]。Favret層の露頭では幅300メートルに達する[11][21]。2つの地層は近接しているが、正確に同じ年代を共有するわけでない。Prida層は中期アニシアン期、Favret層は後期アニシアン期に遡っており[11]、約2億4400万年前と約2億4200万年前にあたる[21]。この時代にFossil Hill部層はパンサラッサ海の東部を形成しており、Benggwigwishingasuchusのような主竜類の存在は沿岸の堆積環境であったことを示唆する[76]。
Fossil Hill部層は海棲爬虫類やアンモナイトおよび他の無脊椎動物が多産しており、表層の海水が曝気されていたことを示唆する[77]。しかし、底生地帯(英語版)はHalobiidae科の二枚貝を除いて動物化石が乏しい。発見された化石から、当該の岩相は当時安定した食物網を持つ遠洋生態系であったことが示唆される。硬骨魚類はほぼ産出しておらず、Favret層でのみ発見されている。発見された魚類について、条鰭類ではサウリクティスや未同定のものがおり[78]、肉鰭類では未同定のシーラカンス科の魚類が多く知られている[21]。Prida層においては多数の軟骨魚類が同定されており、HybodontiformesやSynechodontiformesが知られ、 また分類不確定の板鰓亜綱も産出している[79]。Fossil Hill部層で最も多産する海棲爬虫類は魚鰭類であり、キンボスポンディルスの他に、オムファロサウルスやファラロドン(英語版)およびタラットアルコンが知られる。他の海棲爬虫類はあまり知られておらず、明確に同定されたものとして鰭竜類のAugustasaurusがいる[12][21][80][23]。
ヨーロッパ

C. buschseriの唯一知られている標本は、スイスにおいてGrenzbitumenzoneとしても知られるベザーノ層から記録されている。ベザーノ層はアルプス山脈に分布する地層で、スイスから北部イタリアにかけて広がっており、アニシアン期末からラディニアン期初頭にあたる多数の化石が保存されている[5][59]。ベザーノ層はサン・ジョルジョ山の炭酸塩プラットフォーム(英語版)上に位置する一連の中期三畳紀のユニットの一つであり、層厚は約5 - 16メートルに達する。動物たちが生息しベザーノ層が堆積していた当時、当該地域はテチス海の西部の盆地に位置するラグーンであったと推測されている[81][59]。研究者らの推定によれば、このラグーンの水深は30 - 130メートルに達した[59]。表層付近の海水は酸素が豊富であり、ネクトンやプランクトンが多数生息した[81][82][83]。しかし、ラグーン内の水循環は発達しておらず、水底では典型的な無酸素水(英語版)が広がっていた[82][83]。ラグーンの水底は極めて穏やかであり、その証拠として岩石に細かな葉理が認められること、生物擾乱の証拠がほぼ存在しないことが挙げられる[59]。球果植物や陸棲爬虫類といった陸棲生物の化石が存在することから、当該地域は陸域の付近に位置していたことが示唆される[82]。

ベザーノ層から産出した最も一般的な無脊椎動物の中には、二枚貝類のDaonellaがいる[84]。ベザーノ層からは多数の腹足類も産出しており、主にプランクトンとして、あるいは藻類に付着して生息していたと可能性がある[83]。頭足類ではオウムガイ亜綱(英語版)や鞘形亜綱(英語版)が含まれ、また特にアンモナイトが一般的であった[84]。ベザーノ層産の鞘形類は特に多様性が高いわけでないが、これは鞘形類の遺骸が容易に化石化せず、また数多くの彼らの既知の化石が魚竜の体内の胃内容物として保存されていることによる可能性がある[59][84]。ベザーノ層から産出する節足動物には貝虫や嚢頭類(英語版)およびエビが含まれる。より珍しい無脊椎動物のグループとしては、腕足動物とウニ(棘皮動物)がいる[84][82]。放散虫と海藻も本層から知られているが、他の底生生物と共に別の場所から打ち寄せられた可能性がある[84]。本層では非常に多数の硬骨魚類が記録されており[84][85][86]、特に条鰭類の多様性が高く、豊富な小型種だけでなくサウリクティスのような大型魚類も保存されている[85]。肉鰭類は数が限られており、Rieppelia、Ticinepomis、そしておそらくHolophagusが知られている(いずれもシーラカンス目)[86]。軟骨魚類は多産しないが、主にヒボドゥス目(英語版)に占められる[84][87]。
ネバダ州のFossil Hill部層と異なり、魚竜はベザーノ層において最も多様な海棲爬虫類でなく、ベサノサウルス(英語版)やC. buchseri、ファラロドンおよびミクソサウルスに限られている[88]。これらの魚竜が多産する層準は地層の中部に位置し、当該の年代はラグーンが最も深い時代であった[59]。一方で鰭竜類は当該の海棲爬虫類群集で最大級の規模のグループであり、硬質の食餌を破砕していた板歯目のパラプラコドゥス(英語版)やキアモドゥス(英語版)[89]、また頸部の長いパキプレウロサウルス類やノトサウルス科(英語版)が知られている。パキプレウロサウルス類のOdoiporosaurusはベザーノ層中部から産出しているが、特に多産するSerpianosaurusは魚竜がより希少になりつつあるベザーノ層上部から産出している[84][90]。ノトサウルス科の属にはシルヴェストロサウルス(英語版)やノトサウルスがあり、後者はN. giganteusと、おそらくN. juvenilisと見られる個体を含む[91]。希少であるものの、N. giganteusはC. buschseriのような頂点捕食者であった可能性がある[59]。魚竜と鰭竜類の他では、頸部の長いタニストロフェウスやマクロクネムス(英語版)が知られ[92]、タラットサウルス類(英語版)のAskeptosaurus、Clarazia、Hescheleriaが産出している[93]。
ニッチ分割
Hill Member部層とベザーノ層のいずれにおいても、キンボスポンディルスは多様な魚竜の一つであり、それぞれの属は異なる採餌戦略を採って競争を避けていた(ニッチ分割)と見られる[12][21][59]。キンボスポンディルスと同時代に生息したタラットアルコンは、大型で鋭利な歯を持つことから当時の生態系における唯一の頂点捕食者であったと見られており、キンボスポンディルスはおそらくより小型の海棲爬虫類を攻撃していた[12][21][80][23]。歯の形態と小型さからベサノサウルスは鞘形類の捕食に特化していた可能性が高い。Mixosaurus cornalianusの腹部の内容物は小型の鞘形類と魚類を含んでおり、より大型の類縁種よりも小型の獲物を捕食していたと見られる[59]。より化石の少ないミクソサウルス科であるMixosaurus kuhnschnyderiやファラロドン(英語版)はいずれも破砕に向いた幅広の歯を持つ。M. kuhnschnyderiは鞘形類を摂食したことが知られているが、ファラロドンの持つ大型の歯は獲物の外殻を破砕することに適していた可能性がある[88][21]。オムファロサウルスは、おそらくアンモナイトを磨り潰すことに特化していた[21]。
メディアでの有名な登場
キンボスポンディルスはBBCのドキュフィクション『続 タイムスリップ! 恐竜時代 古代の海へ』(2003年)で、特に様々な三畳紀の海棲爬虫類の登場する章で登場した。登場シーンでは、キンボスポンディルスはプレゼンターのナイジェル・マーヴェンが所持していたタニストロフェウスの尾を奪い、マーヴェンに向かって威嚇した[94]。
注釈
- ^ Leidy (1868)はホロタイプについて僅かに異なる記載を行っており、胴椎の数を4個としている。これは化石の保存に起因するものである[2]。
- ^ Merriam (1908)が記載したC. petrinusの追加標本の1つ[20]。
- ^ Sander et al. (2021)はC. yongorumの95パーセント予測区間に基づいて動物の体サイズを推定した。加えて、研究チームはさらなる2つの推定を行った。予測区間が95パーセント未満の場合、全長12.5メートル体重14.7トン、95パーセントを超過する予測区間の場合、全長25メートル体重135.8トンとなる[12]。推定が95パーセントに近づかないため、これらの推定値はC. youngorumも最大サイズの可能性が高い値とは考えられない[21]。
- ^ これらの観察結果はC. petrinusの特定の標本によって異なるが、UCMP 9950の頭骨では矢状稜の区別がUCMP 9913よりも明確である[11]。
出典
- ^ a b c ティム・ヘインズ、ポール・チェンバーズ 著、椿正晴 訳『よみがえる恐竜・古生物』群馬県立自然史博物館 監修、SBクリエイティブ、65頁。ISBN 4-7973-3547-5。
- ^ a b c d e Joseph Leidy (1868). “Notice of some reptilian remains from Nevada”. Proceedings of the Philadelphia Academy of Sciences 20: 177–178. https://www.biodiversitylibrary.org/page/9768327#page/185/mode/1up.
- ^ a b c Merriam 1902, p. 104.
- ^ Merriam 1908, p. 123.
- ^ a b c d e f g h i j k l m n o P. Martin Sander (1989). “The large ichthyosaur Cymbospondylus buchseri, sp. nov., from the Middle Triassic of Monte San Giorgio (Switzerland), with a survey of the genus in Europe”. Journal of Vertebrate Paleontology 9 (2): 163–173. Bibcode: 1989JVPal...9..163S. doi:10.1080/02724634.1989.10011750. JSTOR 4523251. https://www.researchgate.net/publication/254313617.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y Nadia Fröbisch; P. Martin Sander; Olivier Rieppel (2006). “A new species of Cymbospondylus (Diapsida, Ichthyosauria) from the Middle Triassic of Nevada and a re-evaluation of the skull osteology of the genus”. Zoological Journal of the Linnean Society 147 (4): 515–538. doi:10.1111/j.1096-3642.2006.00225.x.
- ^ a b c d “Cymbospondylus”. Paleofile. 2024年9月7日閲覧。
- ^ Merriam 1908, p. 5-6.
- ^ Merriam 1908, p. 65-70, 104-122.
- ^ a b Michael W. Maisch; Andreas T. Matzke (2004). “Observations on Triassic ichthyosaurs. Part XIII: New data on the cranial osteology of Cymbospondylus petrinus (LEIDY, 1868) from the Middle Triassic Prida Formation of Nevada”. Neues Jahrbuch für Geologie und Paläontologie - Monatshefte 2004 (6): 370–384. doi:10.1127/njgpm/2004/2004/370. https://www.researchgate.net/publication/260273528.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac Nicole Klein; Lars Schmitz; Tanja Wintrich; P. Martin Sander (2020). “A new cymbospondylid ichthyosaur (Ichthyosauria) from the Middle Triassic (Anisian) of the Augusta Mountains, Nevada, USA”. Journal of Systematic Palaeontology 18 (14): 1167–1191. Bibcode: 2020JSPal..18.1167K. doi:10.1080/14772019.2020.1748132. https://www.researchgate.net/publication/340802048.
- ^ a b c d e f g h i j k l m n o p q r P. Martin Sander; Eva Maria Griebeler; Nicole Klein; Jorge Velez Juarbe; Tanja Wintrich; Liam J. Revell; Lars Schmitz (2021). “Early giant reveals faster evolution of large body size in ichthyosaurs than in cetaceans”. Science 374 (6575): eabf5787. doi:10.1126/science.abf5787. PMID 34941418.
- ^ Merriam 1908, p. 123-124.
- ^ McGowan & Motani 2003, p. 65-66.
- ^ Wolniewicz 2017, p. 123.
- ^ International Commission on Zoological Nomenclature (2012年). “Article 8. What constitutes published work”. International Code of Zoological Nomenclature. July 16, 2021閲覧。
- ^ Mike Taylor (June 8, 2010). “Notes on Early Mesozoic Theropods and the future of zoological nomenclature”. Sauropod Vertebra Picture of the Week. March 9, 2021時点のオリジナルよりアーカイブ。2024年9月7日閲覧。
- ^ Bernhard Peyer (1944). “Die Reptilien vom Monte San Giorgio [The reptiles of Monte San Giorgio]” (ドイツ語). Neujahrsblau der Naturforschcnden Gesellschaft in Zürich 146: 1–95.
- ^ Emil Kuhn-Schnyder (1964). “Die Wirbellierfauna der Tessiner Kalkalpen [The vertebrate fauna of the Ticino Limestone Alps]” (ドイツ語). Geologische Rundschau 53: 393–412. doi:10.1007/BF02040759.
- ^ Merriam 1908, p. 105-110.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y P. Martin Sander; Eva Maria Griebeler; Lars Schmitz (2021). “Supplementary Materials for Early giant reveals faster evolution of large body size in ichthyosaurs than in cetaceans”. Science 374 (6575): eabf5787. doi:10.1126/science.abf5787. PMID 34941418. https://www.science.org/action/downloadSupplement?doi=10.1126%2Fscience.abf5787&file=science.abf5787_sm.pdf.
- ^ Wolniewicz 2017, p. 149.
- ^ a b c Nadia B. Fröbisch; Jörg Fröbisch; P. Martin Sander; Lars Schmitz; Olivier Rieppel (2013). “Supporting Information”. Proceedings of the National Academy of Sciences of the United States of America 110 (4): 1393–1397. Bibcode: 2013PNAS..110.1393F. doi:10.1073/pnas.1216750110. PMC 3557033. PMID 23297200. https://www.pnas.org/action/downloadSupplement?doi=10.1073%2Fpnas.1216750110&file=pnas.201216750si.pdf.
- ^ Alexandra Houssaye; Yasuhisa Nakajima; P. Martin Sander (2018). “Structural, functional, and physiological signals in ichthyosaur vertebral centrum microanatomy and histology”. Geodiversitas 40 (2): 161–170. doi:10.5252/geodiversitas2018v40a7. https://www.researchgate.net/publication/324484636.
- ^ Lene Liebe Delsett; Nicholas D. Pyenson (2021). “Early and fast rise of Mesozoic ocean giants”. Science 374 (6575): 1554–1555. Bibcode: 2021Sci...374.1554D. doi:10.1126/science.abm3751. PMID 34941421.
- ^ a b c Judy A. Massare; Jack M. Callaway (1994). “Cymbospondylus (Ichthyosauria: Shastasauridae) from the Lower Triassic Thaynes Formation of southeastern Idaho”. Journal of Vertebrate Paleontology 14 (1): 139–141. Bibcode: 1994JVPal..14..139M. doi:10.1080/02724634.1994.10011545. JSTOR 4523552.
- ^ a b c Victoria S. Engelschiøn; Lene L. Delsett; Aubrey J. Roberts; Jørn H. Hurum (2018). “Large-sized ichthyosaurs from the Lower Saurian niveau of the Vikinghøgda Formation (Early Triassic), Marmierfjellet, Spitsbergen”. Norwegian Journal of Geology 98 (2): 239–265. doi:10.17850/njg98-2-05. hdl:10852/71102.
- ^ a b c d e f g h i j P. Martin Sander; René Dederichs; Tanja Schaaf; Eva Maria Griebeler (2024). “Cymbospondylus (Ichthyopterygia) from the Early Triassic of Svalbard and the early evolution of large body size in ichthyosaurs”. PalZ 98 (2): 275–290. Bibcode: 2024PalZ...98..275S. doi:10.1007/s12542-023-00677-3.
- ^ Emil Kuhn-Schnyder (1980). “Über Reste eines großen Ichthyosauriers aus den Buchensteiner Schichten (ladinische Stufe der Trias) der Seceda (NE St. Ulrich/Ortisei, Prov. Bozen, Italien) [About the remains of a large ichthyosaur from the Buchenstein layers (Ladin stage of the Triassic) of the Seceda (NE St. Ulrich/Ortisei, Prov. Bozen, Italy)]” (ドイツ語). Annalen des Naturhistorischen Museums in Wien 83: 231–244. https://www.zobodat.at/pdf/ANNA_83_0231-0244.pdf.
- ^ a b c d Marco Balini; Silvio C. Renesto (2012). “Cymbospondylus vertebrae (Ichthyosauria, Shastasauridae) from the Upper Anisian Prezzo Limestone (Middle Triassic, Southern Alps) with an overview of the chronostratigraphic distribution of the group”. Rivista Italiana di Paleontologia e Stratigrafia 118 (1): 155–172. doi:10.13130/2039-4942/5996.
- ^ P. Martin Sander (1992). “Cymbospondylus (Shastasauridae: Ichthyosauria) from the Middle Triassic of Spitsbergen: filling a paleobiogeographic gap”. Journal of Paleontology 66 (2): 332–337. Bibcode: 1992JPal...66..332S. doi:10.1017/S0022336000033825. JSTOR 1305917. https://www.researchgate.net/publication/260391769.
- ^ Olivier Rieppel; Fabio Marco Dalla Vecchia (2001). “Marine reptiles from the Triassic of the Tre Venezie Area, Northeastern Italy”. Fieldiana, Geology. New Series 44: 1–25. https://www.biodiversitylibrary.org/item/25238#page/3/mode/1up.
- ^ a b Erin E. Maxwell; Benjamin P. Kear (2013). “Triassic ichthyopterygian assemblages of the Svalbard archipelago: A reassessment of taxonomy and distribution”. GFF 135 (1): 85–94. Bibcode: 2013GFF...135...85M. doi:10.1080/11035897.2012.759145. https://www.researchgate.net/publication/263214147.
- ^ Merriam 1902, p. 106-107.
- ^ Merriam 1908, p. 104.
- ^ John W. Hulke (1873). “Memorandum on some fossil vertebrate remains collected by the Swedish expeditions to Spitzbergen in 1864 and 1868”. Bihang till Kongliga Svenska Vetenskaps-Akademiens Handlingar 1 (9): 1–11. https://www.biodiversitylibrary.org/item/49831#page/419/mode/1up.
- ^ Nikolai N. Yakowlew (1902). “Neue funde von Trias-Sauriern auf Spitzbergen [New discoveries of Triassic reptiles on Spitsbergen]” (ドイツ語). Verhandlungen der Russisch-Kaiserlichen Mineralogischen Gesellschaft zu St. Petersburg 40: 179–202.
- ^ Merriam 1908, p. 149-150.
- ^ Carl Wiman (1910). “Ichthyosaurier aus der Trias Spitzbergens [Ichthyosaurs from the Triassic of Spitsbergen]” (ドイツ語). Bulletin of the Geological Institutions of the University of Uppsala 10: 124–148. https://paleoarchive.com/literature/Wiman1910-IchthyosaurierTriasSpitzbergens.pdf.
- ^ McGowan & Motani 2003, p. 128.
- ^ Merriam 1908, p. 124-127.
- ^ McGowan & Motani 2003, p. 125.
- ^ Merriam 1908, p. 150-152.
- ^ John C. Merriam (1911). “Notes on the relationships of the marine saurian fauna described from the Triassic of Spitzbergen by Wiman”. University of California Publications. Bulletin of the Department of Geology 6 (13): 317–327. https://www.biodiversitylibrary.org/item/139555#page/427/mode/1up.
- ^ Elizabeth L. Nicholls; Donald B. Brinkman; Jack M. Callaway (1999). “New Material of Phalarodon (Reptilia: Ichthyosauria) from the Triassic of British Columbia and its Bearing on the Interrelationships of Mixosaurs”. Palaeontographica Abteilung A 252 (1): 1–22. doi:10.1127/pala/252/1998/1. https://www.researchgate.net/publication/286357136.
- ^ Lars Schmitz (2005). “The taxonomic status of Mixosaurus nordenskioeldii (Ichthyosauria)”. Journal of Vertebrate Paleontology 25 (4): 983–985. doi:10.1671/0272-4634(2005)025[0983:TTSOMN]2.0.CO;2. JSTOR 4524525.
- ^ Friedrich von Huene (1916). “Beiträge zur Kenntnis der Ichthyosaurier im deutschen Muschelkalk [Contributions to the knowledge of ichthyosaurs in the German Muschelkalk]” (ドイツ語). Palaeontographica 68: 1–68. https://www.biodiversitylibrary.org/item/110906#page/13/mode/1up.
- ^ Von F. Broili (1916). “Einige Bemerkungen über die Mixosauridae [Some remarks about the Mixosauridae]” (ドイツ語). Anatomischer Anzeiger 49: 474–494. https://www.biodiversitylibrary.org/item/47658#page/490/mode/1up.
- ^ Jack M. Callaway; Judith A. Massare (1989). “Geographic and stratigraphic distribution of the Triassic Ichthyosauria (Reptilia; Diapsida)”. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 178: 37–58. doi:10.1127/njgpa/178/1989/37.
- ^ McGowan & Motani 2003, p. 129.
- ^ Chun Li; Hai-Lu You (2002). “Cymbospondylus from the Upper Triassic of Guizhou, China” (zh, en). Vertebrata PalAsiatica 40: 9–16. http://www.ivpp.cas.cn/cbw/gjzdwxb/xbwzxz/200901/W020090813372039067086.pdf.
- ^ Qing-Hua Shang; Chun Li (2009). “On the occurrence of the ichthyosaur Shastasaurus in the Guanling biota (Late Triassic), Guizhou, China” (zh, en). Vertebrata PalAsiatica 47 (3): 178–193. http://www.ivpp.cas.cn/cbw/gjzdwxb/xbwzxz/200909/P020090917531791680790.pdf.
- ^ Maisch 2010, p. 163.
- ^ P. Martin Sander; Xiaohong Chen; Long Cheng; Xiaofeng Wang (2011). “Short-Snouted Toothless Ichthyosaur from China Suggests Late Triassic Diversification of Suction Feeding Ichthyosaurs”. PLOS ONE 6 (5): e19480. Bibcode: 2011PLoSO...619480S. doi:10.1371/journal.pone.0019480. PMC 3100301. PMID 21625429. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3100301/.
- ^ Qing-Hua Shang; Chun Li (2013). “On the sexual dimorphism of Shastasaurus tangae (Reptilia: Ichthyosauria) from the Triassic Guanling Biota, China” (zh, en). Vertebrata PalAsiatica 51 (4): 253–264. http://www.ivpp.cas.cn/cbw/gjzdwxb/xbwzxz/201312/P020131205319316919512.pdf.
- ^ Cheng Ji; Da-Yong Jiang; Ryosuke Motani; Olivier Rieppel; Wei-Cheng Hao; Zuo-Yu Sun (2016). “Phylogeny of the Ichthyopterygia incorporating recent discoveries from South China”. Journal of Vertebrate Paleontology 36 (1): e1025956. Bibcode: 2016JVPal..36E5956J. doi:10.1080/02724634.2015.1025956. https://www.researchgate.net/publication/283840343.
- ^ Benjamin C. Moon (2019). “A new phylogeny of ichthyosaurs (Reptilia: Diapsida)”. Journal of Systematic Palaeontology 17 (2): 129–155. Bibcode: 2019JSPal..17..129M. doi:10.1080/14772019.2017.1394922. hdl:1983/463e9f78-10b7-4262-9643-0454b4aa7763. https://core.ac.uk/download/pdf/146503337.pdf.
- ^ Da-Yong Jiang; Ryosuke Motani; Andrea Tintori; Olivier Rieppel; Cheng Ji; Min Zhou; Xue Wang; Hao Lu et al. (2020). “Evidence supporting predation of 4-m marine reptile by Triassic megapredator”. iScience 23 (9): 101347. Bibcode: 2020iSci...23j1347J. doi:10.1016/j.isci.2020.101347. PMC 7520894. PMID 32822565. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7520894/.
- ^ a b c d e f g h i j k l m n Gabriele Bindellini; Andrzej S. Wolniewicz; Feiko Miedema; Torsten M. Scheyer; Cristiano Dal Sasso (2021). “Cranial anatomy of Besanosaurus leptorhynchus Dal Sasso & Pinna, 1996 (Reptilia: Ichthyosauria) from the Middle Triassic Besano Formation of Monte San Giorgio, Italy/Switzerland: taxonomic and palaeobiological implications”. PeerJ 9: e11179. doi:10.7717/peerj.11179. PMC 8106916. PMID 33996277. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8106916/.
- ^ a b Ryan Marek (2015). “Fossil Focus: Ichthyosaurs”. Palaeontology Online 5: 8. オリジナルの18 January 2021時点におけるアーカイブ。. https://web.archive.org/web/20210118111725/https://www.palaeontologyonline.com/articles/2015/fossil-focus-ichthyosaurs/ 13 June 2020閲覧。.
- ^ P. Martin Sander (2000). “Ichthyosauria: their diversity, distribution, and phylogeny”. Paläontologische Zeitschrift 74 (1): 1–35. Bibcode: 2000PalZ...74....1S. doi:10.1007/BF02987949. https://www.researchgate.net/publication/226305457.
- ^ Ryosuke Motani (2005). “Evolution of Fish-Shaped Reptiles (reptilia: Ichthyopterygia) in Their Physical Environments and Constraints”. Annual Review of Earth and Planetary Sciences 33: 395–420. Bibcode: 2005AREPS..33..395M. doi:10.1146/annurev.earth.33.092203.122707. オリジナルの2018-12-23時点におけるアーカイブ。. https://web.archive.org/web/20181223015949/http://mygeologypage.ucdavis.edu/motani//pdf/Motani2005AnnRev.pdf.
- ^ Judith M. Pardo-Pérez; Benjamin P. Kear; Erin E. Maxwell (2020). “Skeletal pathologies track body plan evolution in ichthyosaurs”. Scientific Reports 10 (1): 4206. Bibcode: 2020NatSR..10.4206P. doi:10.1038/s41598-020-61070-7. PMC 7060314. PMID 32144303. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7060314/.
- ^ McGowan & Motani 2003, p. 66.
- ^ a b c Merriam 1908, p. 105.
- ^ Dean R. Lomax; Paul de la Salle; Marcello Perillo; Justin Reynolds; Ruby Reynolds; James F. Waldron (2024). “The last giants: New evidence for giant Late Triassic (Rhaetian) ichthyosaurs from the UK”. PLOS ONE 19 (4): e0300289. Bibcode: 2024PLoSO..1900289L. doi:10.1371/journal.pone.0300289. PMC 11023487. PMID 38630678. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC11023487/.
- ^ Merriam 1908, p. 106.
- ^ Merriam 1908, p. 107.
- ^ a b Merriam 1908, p. 110.
- ^ Ryosuke Motani (1997), “Temporal and Spatial Distribution of Tooth Implantations in Ichthyosaurs”, in Callaway, Jack M.; Nicholls, Elizabeth L., Ancient Marine Reptiles, San Diego: Academic Press, pp. 81–103, doi:10.1016/B978-012155210-7/50007-7, ISBN 978-0-12-155210-7
- ^ Merriam 1908, p. 111.
- ^ Maisch 2010, p. 155.
- ^ Ryosuke Motani (1999). “Phylogeny of the Ichthyopterygia”. Journal of Vertebrate Paleontology 19 (3): 472–495. Bibcode: 1999JVPal..19..473M. doi:10.1080/02724634.1999.10011160. JSTOR 4524011. オリジナルの2016-03-05時点におけるアーカイブ。. https://web.archive.org/web/20160305104153/http://mygeologypage.ucdavis.edu/motani/pdf/Motani1999c.pdf 2011年10月19日閲覧。.
- ^ Massare, J.A.; Callaway, JM (1990). “The affinities and ecology of Triassic ichthyosaurs”. Geological Society of America Bulletin 102 (4): 409–416. Bibcode: 1990GSAB..102..409M. doi:10.1130/0016-7606(1990)102<0409:TAAEOT>2.3.CO;2. https://pubs.geoscienceworld.org/gsa/gsabulletin/article-abstract/102/4/409/182405/The-affinities-and-ecology-of-Triassic.
- ^ Nichols & Silberling 1977, p. 20.
- ^ Nathan D. Smith; Nicole Klein; P. Martin Sander; Lars Schmitz (2024). “A new pseudosuchian from the Favret Formation of Nevada reveals that archosauriforms occupied coastal regions globally during the Middle Triassic”. Biology Letters 20 (7): 20240136. doi:10.1098/rsbl.2024.0136. PMC 11286145. PMID 38982977. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC11286145/.
- ^ Nichols & Silberling 1977, p. 18.
- ^ P. Martin Sander; Olivier C. Rieppel; H. Bucher (1994). “New Marine Vertebrate Fauna from the Middle Triassic of Nevada”. Journal of Paleontology 68 (3): 676–680. Bibcode: 1994JPal...68..676S. doi:10.1017/S0022336000026020. JSTOR 1306213.
- ^ Gilles Cuny; Olivier Rieppel; P. Martin Sander (2001). “The shark fauna from the Middle Triassic (Anisian) of North-Western Nevada”. Zoological Journal of the Linnean Society 133 (3): 285–301. doi:10.1111/j.1096-3642.2001.tb00627.x.
- ^ a b Nadia B. Fröbisch; Jörg Fröbisch; P. Martin Sander; Lars Schmitz; Olivier Rieppel (2013). “Macropredatory ichthyosaur from the Middle Triassic and the origin of modern trophic networks”. Proceedings of the National Academy of Sciences of the United States of America 110 (4): 1393–1397. Bibcode: 2013PNAS..110.1393F. doi:10.1073/pnas.1216750110. PMC 3557033. PMID 23297200. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3557033/.
- ^ a b Silvio Renesto; Cristiano Dal Sasso; Fabio Fogliazza; Cinzia Ragni (2020). “New findings reveal that the Middle Triassic ichthyosaur Mixosaurus cornalianus is the oldest amniote with a dorsal fin”. Acta Palaeontologica Polonica 65 (3): 511–522. doi:10.4202/app.00731.2020.
- ^ a b c d Heinz Furrer (1995). “The Kalkschieferzone (Upper Meride Limestone, Ladinian) near Meride (Canton Ticino, Southern Switzerland) and the evolution of a Middle Triassic intraplatform basin”. Eclogae Geologicae Helvetiae 88 (3): 827–852. https://www.e-periodica.ch/cntmng?pid=egh-001:1995:88::1037.
- ^ a b c Vittorio Pieroni; Heinz Furrer (2020). “Middle Triassic gastropods from the Besano Formation of Monte San Giorgio, Switzerland”. Swiss Journal of Palaeontology 139 (1): 2. Bibcode: 2020SwJP..139....2P. doi:10.1186/s13358-019-00201-8. ISSN 1664-2384.
- ^ a b c d e f g h Hans-Joachim Röhl; Annette Schmid-Röhl; Heinz Furrer; Andreas Frimmel; Wolfgang Oschmann; Lorenz Schwark (2001). “Microfacies, geochemistry and palaeoecology of the Middle Triassic Grenzbitumenzone from Monte San Giorgio (Canton Ticino, Switzerland)”. Geologia Insubrica 6: 1–13. https://www.researchgate.net/publication/266096095.
- ^ a b Carlo Romano (2021). “A Hiatus Obscures the Early Evolution of Modern Lineages of Bony Fishes”. Frontiers in Earth Science 8: 618853. doi:10.3389/feart.2020.618853. ISSN 2296-6463.
- ^ a b Christophe Ferrante; Lionel Cavin (2023). “Early Mesozoic burst of morphological disparity in the slow-evolving coelacanth fish lineage”. Scientific Reports 13 (1): 11356. Bibcode: 2023NatSR..1311356F. doi:10.1038/s41598-023-37849-9. PMC 10345187. PMID 37443368. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10345187/.
- ^ John G. Maisey (2011). “The braincase of the Middle Triassic shark Acronemus tuberculatus (Bassani, 1886)”. Palaeontology 54 (2): 417–428. Bibcode: 2011Palgy..54..417M. doi:10.1111/j.1475-4983.2011.01035.x.
- ^ a b Winand Brinkmann (2004). “Mixosaurier (Reptilia, Ichthyosaurier) mit Quetschzähnen aus der Grenzbitumenzone (Mitteltrias) des Monte San Giorgio (Schweiz, Kanton Tessin) [Mixosaurs (Reptilia, Ichthyosauria) with crushing teeth from the Grenzbitumenzone (Middle Triassic) of Monte San Giorgio (Switzerland, Canton of Ticino)]” (en, de). Schweizerische Paläontologische Abhandlungen 124: 1–86. https://www.researchgate.net/publication/308170386.
- ^ Michael W. Maisch (2020). “The evolution of the temporal region of placodonts (Diapsida: Placodontia) – a problematic issue of cranial osteology in fossil marine reptiles”. Palaeodiversity 13 (1): 57–68. doi:10.18476/pale.v13.a6.
- ^ Nicole Klein; Heinz Furrer; Iris Ehrbar; Marta Torres Ladeira; Henning Richter; Torsten M. Scheyer (2022). “A new pachypleurosaur from the Early Ladinian Prosanto Formation in the Eastern Alps of Switzerland”. Swiss Journal of Palaeontology 141 (1): 12. Bibcode: 2022SwJP..141...12K. doi:10.1186/s13358-022-00254-2. PMC 9276568. PMID 35844249. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9276568/.
- ^ Silvio Renesto (2010). “A new specimen of Nothosaurus from the latest Anisian (Middle Triassic) Besano formation (Grenzbitumenzone) of Italy”. Rivista Italiana di Paleontologia e Stratigrafia 116 (2): 145–160. doi:10.13130/2039-4942/5946.
- ^ Stephan N.F. Spiekman; James M. Neenan; Nicholas C. Fraser; Vincent Fernandez; Olivier Rieppel; Stefania Nosotti; Torsten M. Scheyer (2020). “The cranial morphology of Tanystropheus hydroides (Tanystropheidae, Archosauromorpha) as revealed by synchrotron microtomography”. PeerJ 8: e10299. doi:10.7717/peerj.10299. PMC 7682440. PMID 33240633. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7682440/.
- ^ Nicole Klein; P. Martin Sander; Jun Liu; Patrick S. Druckenmiller; Eric T. Metz; Neil P. Kelley; Torsten M. Scheyer (2023). “Comparative bone histology of two thalattosaurians (Diapsida: Thalattosauria): Askeptosaurus italicus from the Alpine Triassic (Middle Triassic) and a Thalattosauroidea indet. from the Carnian of Oregon (Late Triassic)”. Swiss Journal of Palaeontology 142 (1): 15. Bibcode: 2023SwJP..142...15K. doi:10.1186/s13358-023-00277-3. PMC 10432342. PMID 37601161. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10432342/.
- ^ Nigel Marven; Jasper James (2004). Chased by Sea Monsters: Prehistoric Predators of the Deep. New York: DK. pp. 74–79. ISBN 978-0-7566-0375-5. OCLC 1391299004. https://archive.org/details/chasedbyseamonst0000marv/page/74/mode/2up
参考文献
- Merriam, John C. (1902). “Triassic Ichthyopterygia from California and Nevada”. University of California Publications. Bulletin of the Department of Geology 3: 63–108. https://www.biodiversitylibrary.org/item/148596#page/91/mode/1up.
- Merriam, John C. (1908). Triassic Ichthyosauria, with special reference to the American forms. Memoirs of the University of California. 1. Berkeley, California: The University Press. pp. 1–196. OCLC 457714430. https://www.biodiversitylibrary.org/item/60829#page/9/mode/1up
- Nichols, Kathryn M.; Silberling, Norman J. (1977). Stratigraphy and depositional history of the Star Peak Group (Triassic), northwestern Nevada. 175-178. Boulder: Geological Society of America, Special Papers. pp. 1–73. doi:10.1130/SPE178-p1. ISBN 978-0-813-72178-1. https://books.google.com/books?id=ojSgyTAPuJgC
- Maisch, Michael W.; Matzke, Andreas T. (2000). “The Ichthyosauria”. Stuttgarter Beiträge zur Naturkunde, Serie B 298: 1–159. https://www.biodiversitylibrary.org/item/278993#page/350/mode/2up.
- McGowan, Christopher; Motani, Ryosuke (2003). Handbook of Paleoherpetology Part 8: Ichthyopterygia. 1. Munich: Verlag Dr. Friedrich Pfeil. pp. 1–175. ISBN 978-3-899-37007-2. OCLC 469848769. http://beeld.teylersmuseum.nl/Digital_Library/Emags/Saur_028/page_1.html
- Maisch, Michael W. (2010). “Phylogeny, systematics, and origin of the Ichthyosauria – the state of the art”. Palaeodiversity 3: 151–214. http://www.palaeodiversity.org/pdf/03/Palaeodiversity_Bd3_Maisch.pdf.
- Wolniewicz, Andrzej S. (2017). The Anatomy, Taxonomy and Systematics of Middle Triassic–Early Jurassic Ichthyosaurs (Reptilia: Ichthyopterygia) and the Phylogeny of Ichthyopterygia (PhD). University of Oxford.









